На основании анализа родословной животного можно качественно оценить степень инбридинга, которая выражается насыщенностью родословной общими предками (кличками) по материнской и отцовской линиям.

По Шапоружу, римскими цифрами обозначаются поколения (ряд предков), считая родительское за первое, дедовское за второе, и т.д. Если общий предок повторяется и в материнской, и в отцовской части родословной, то сначала ставится его встречаемость в материнской родословной, а затем, через чёрточку, - в отцовской. Тогда скрещивание сын х мать будет обозначаться как I-II (рис. 2,а), отец х дочь как II-I (рис. 2,6), брат х сестра как II-II (рис. 2,в, г), дед х внучка как III—I (рис. 2,д), двоюродные брат х сестра как III- III (рис. 2,е), и т. п.
- Инбридинг типа отец х дочь (II-I), сын х мать (I-II) и брат х сестра (II-II) обозначается как очень тесный, или кровосмешение;
- близкородственное скрещивание типа II-III или III-II - это тесный инбридинг;
- а скрещивание III-IV, IV-III, IV-IV, II-V и т. п. - умеренный инбридинг.
Ввиду отсутствия ощутимых эффектов более отдалённые родственные скрещивания на практике можно не рассматривать как истинный инбридинг. Владельцы высококлассных породистых кошек, взяв в руки родословную кошки, могут сами оценить, как часто и в какой степени использовался инбридинг заводчиками, участвовавшими в выведении этого питомца.
Для количественного учёта степени инбридинга в родословной животного и тем самым определения меры его гомозиготности английский генетик С. Райт ввёл понятие коэффициента инбридинга (F). Этот коэффициент вычисляется следующим образом.
Сначала считается количество поколений (стрелок - таких, как на рис. 2) от общего предка А отца и матери до отца данного животного (р), затем количество поколений (стрелок) от этого же предка А до матери (m).
Эти числа суммируются, и к сумме прибавляется единица: n = p + m + 1.
Коэффициент инбридинга анализируемого животного (пробанда X) по этому предку равен 1/2 в степени n : FА = (1/2)n.
Если у отца и матери оказывается несколько общих предков, то коэффициенты суммируются: Fx = ∑Fi, =∑(1/2)n, где i - общие предки А, В, С и т. д.
 Для примера рассчитаем коэффициент инбридинга животного (пробанда X), полученного от скрещивания родных брата и сестры (II-II, рис. 2,в). В родословной расстояние (путь) от деда до отца - 1 поколение, от деда до матери - 1 поколение, следовательно,
Для примера рассчитаем коэффициент инбридинга животного (пробанда X), полученного от скрещивания родных брата и сестры (II-II, рис. 2,в). В родословной расстояние (путь) от деда до отца - 1 поколение, от деда до матери - 1 поколение, следовательно,
n=1+1+1=3, а значит FB = (1/2)3 = 1/8.
Но у них есть ещё один общий предок - их бабушка; от неё до отца - 1 поколение, от неё же до матери - 1 поколение, а стало быть FA будет тоже 1/8. Общий коэффициент инбридинга для X получается:
Fх = FA + FB = 1/8+1/8=1/4, или 0,25.
Другой пример - скрещивание деда с внучкой (III—I рис. 2,д).
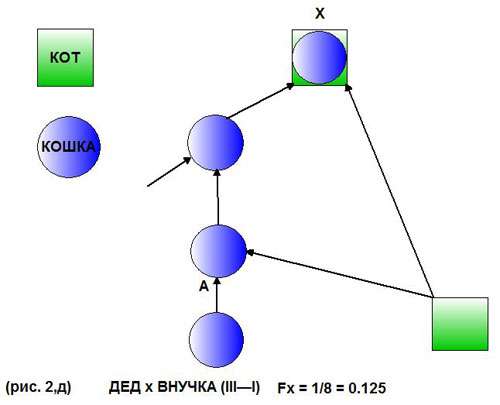 Здесь у потомства по отцовской линии поколений (стрелок) нет, т.е. 0 поколений, по материнской линии - 2 поколения, откуда
Здесь у потомства по отцовской линии поколений (стрелок) нет, т.е. 0 поколений, по материнской линии - 2 поколения, откуда
n = 0 + 2 + 1 = 3, и тогда FX = (1/2)3 = 1/8, или 0,125.
 Наконец, пример со скрещиванием двоюродных брата с сестрой (III- III, рис. 2,е). Общие предки их отпрыска - прадедушка и прабабушка. От прадедушки до отца - 2 поколения, от него же до матери - 2 поколения, т.е.
Наконец, пример со скрещиванием двоюродных брата с сестрой (III- III, рис. 2,е). Общие предки их отпрыска - прадедушка и прабабушка. От прадедушки до отца - 2 поколения, от него же до матери - 2 поколения, т.е.
n = 2 + 2 + 1 = 5, и соответственно FB = (1/2)5 = 1/32.
То же получается при расчёте коэффициента инбридинга на другого общего предка - прабабушку: FA = (1/2)5 = 1/32.
Следовательно, общий коэффициент инбридинга FХ = FA + FB = 1/32 + 1/32 = 1/16, или 0,0625.
Из имеющихся родословных каждый заводчик или владелец породистого животного может рассчитать коэффициент инбридинга своих питомцев или их родителей и прародителей - если в их производстве использовался инбридинг.
Как уже говорилось, этот коэффициент является показателем степени чистоты (гомозиготности) линий, участвующих в родословной, а также указанием на возможную препотентность соответствующих производителей и/или производительниц (см. ниже). Чем выше коэффициент инбридинга животного, тем больше его генов находится в гомозиготном состоянии. Естественно, как следует из формулы, коэффициент инбридинга не может превышать единицы.
ИНОРОДНАЯ ДЕПРЕССИЯ И ГЕТЕРОЗИС
Почему же в человеческом обществе в большинстве цивилизаций существует либо прямой запрет на кровосмешение, либо осуждение его? Более того, почему в дикой природе тоже всё приспособлено так, чтобы по возможности уйти от инбридинга и гомозиготности?
Главная причина в том, что и дикой природе, и человеческому обществу для успешного выживания и приспособляемости к меняющимся условиям окружающей среды (в том числе социальной и экономической в случае человеческого общества) необходимо разнообразие генотипов и особей в популяции.
В разнообразном сообществе всегда найдутся особи, лучше всех приспособленные к данной конкретной ситуации; однако ситуация меняется - и другие особи, с другими признаками выплывают на поверхность. Однообразие популяции, хотя и хорошо приспособленной к данным конкретным условиям её существования, обрекает биологический вид на вымирание при изменении условий существования.
Таким образом, стремление к созданию разнообразия необходимо не отдельным особям, а всему виду для удовлетворения различным и меняющимся требованиям окружающей среды и исторического процесса.
Но существует также и прямая опасность инбридинга для отдельных особей. Дело в том, что как в процессе индивидуальной жизни животного (и человека), так и на протяжении поколений время от времени происходят мутации - изменения генов.
Большая часть мутаций - вредные, и если бы они немедленно проявлялись, то приводили бы к смерти (летальные мутации), уродствам, ненормальностям поведения, несоответствиям окружающей среде, и т.п. Однако большая часть мутаций в то же время имеет рецессивный характер, т.е. в комбинации с нормальным парным геном, благодаря двойному набору генов, не проявляется в гетерозиготном состоянии. Груз таких вредных, но не проявляемых, скрытых мутаций имеется у каждого гетерозиготного индивидуума, включая каждого человека.
Инбридинг создаёт гомозиготность, в том числе гомозиготность по вредным генам, и, следовательно, ведёт к их фенотипическому проявлению в потомстве - внутриутробной или ранней смерти, уродствам, пониженной жизнеспособности и т.п. Это явление называется инбредной депрессией.
Зададим обратный вопрос - почему же, несмотря на опасность инбредной депрессии, селекционеры, выводящие и совершенствующие домашних животных, столь часто прибегают к инбридингу и не боятся его?
Ответ прост: селекционеры не стремятся обеспечить максимальную выживаемость, жизнестойкость и высокие качества всех особей потомства, а ведут отбор лучших, выбраковывая (например, кастрируя) дефектных, слабых и просто с нежелательными признаками. Тем самым при инбридинге, когда происходит разложение генотипа на гомозиготные линии, вредные рецессивные гены проявляются и убираются из дальнейшего разведения, а все "хорошие" гены и их комбинации закрепляются в гомозиготном состоянии и оставляются для дальнейших поколений.
Таким образом, инбридинг в руках селекционера - это способ удаления из генотипа груза вредных мутаций, с одной стороны, и создания необходимых комбинаций ценных генов и признаков, с другой. Кроме того, благодаря гомозиготности создаётся необходимая для выведения любой новой породы однородность, отвечающая требованиям этой породы, и соответствующие ценные признаки оказываются закреплёнными в потомстве. Следует отметить, что по мере инбридинга и отбора лучших, вследствие избавления от груза вредных скрытых мутаций, опасность инбредной депрессии при близкородственном скрещивании отобранных инбредных животных уменьшается.
При скрещивании между представителями разных независимых инбредных линий может наблюдаться явление, в известном смысле противоположное депрессии - гетерозис, называемый также "гибридной силой". Такие гибриды двух инбредных животных обычно обнаруживают повышенную жизнеспособность, рост, плодовитость и т. п. Это происходит потому, что при скрещивании между двумя инбредными линиями, отобранными и потому сходными по типичным признакам породы, сохраняется в потомстве гомозиготность по этим отобранным признакам, тогда как оставшиеся не выбракованными вредные мутации переводятся в гетерозиготное состояние и тем самым становятся рецессивными, т.е. не проявляются.
Авторы: М.Асланян (профессор кафедры генетики и селекции, доктор биологических наук); А.Спирин, (профессор кафедры молекулярной биологии, доктор биологических наук, действительный член Российской академии наук). Статья опубликована в журнале Друг кошек 1996-06



